

La ceruloplasmina (CP) es una metaloproteína con actividad enzimática que presenta la capacidad de transportar el 95% del cobre plasmático. Además, participa del metabolismo del hierro. Su función ferroxidasa posibilita la inactivación de los radicales libres y otros marcadores de estrés oxidativo. Se la considera un biomarcador emergente de procesos inflamatorios, por lo tanto los niveles elevados de CP sérica se asocian con síndrome coronario agudo y enfermedades neoplásicas. El conocimiento de su estructura molecular permitió distinguir diferentes sitios de unión a ligandos. Éstos se hallan relacionados a la multiplicidad de funciones que puede ejercer de acuerdo al contexto biológico en el que se encuentre, por lo cual recibe el nombre de proteína multifuncional. En el caso particular de ausencia o deficiencia genética de la CP se observó depósito de hierro en tejido cerebral, hepático, pancreático, etc. Con respecto a su utilidad terapéutica se ha propuesto como molécula blanco en distintos tipos de cánceres.
Palabras clave: Ceruloplasmina humana; Proteínas multifuncionales; Reactantes de fase aguda; Ferroxidasa; Multicobre-oxidasa
Human ceruloplasmin: a multifunctional protein
Ceruloplasmin (CP) is a metalloprotein with enzymatic activity, capable of transporting 95% of plasma copper. It is also involved in iron metabolism. Its ferroxidase function enables the inactivation of free radicals and other markers of oxidative stress, and it is considered an emerging biomarker of inflammatory processes. Thus, elevated serum PC levels are associated with acute coronary syndrome and neoplastic diseases. Knowledge of its molecular structure has made it possible to distinguish different ligand binding sites. These are related to the multiplicity of functions it can exert according to the biological context in which it is present, which is why it is called moonlighting protein. In the particular case of absence or genetic deficiency of CP, iron deposition was observed in the brain, liver, pancreatic tissue, etc. Regarding its therapeutic usefulness, it has been proposed as a target molecule in different types of cancers.
Keywords: Human ceruloplasmin; Moonlighting protein; Acute phase reactant; Ferroxidase; Multicopper oxidase
Ceruloplasmina humana: uma proteína multifuncional
Resumo
A ceruloplasmina (CP) é uma metaloproteína com atividade enzimática, que tem a capacidade de transportar 95% do cobre plasmático. Também está envolvida no metabolismo do ferro. A sua função ferroxidase permite a inativação de radicais livres e de outros marcadores de estresse oxidativo. É considerado um biomarcador emergente de processos inflamatórios. Assim, os níveis elevados de CP sérica estão associados à síndrome coronariana aguda e à doenças neoplásicas. O conhecimento da sua estrutura molecular permitiu distinguir diferentes locais de ligação dos ligandos. Eles estão relacionados com a multiplicidade de funções que pode exercer de acordó com o contexto biológico no qual se encontre. Por esse motivo, recebe o nome de proteína multifuncional. No caso particular da ausência ou deficiência genética da CP, foram observados depósitos de ferro em tecido cerebral, hepático, pancreático, etc. No que se refere à sua utilidade terapêutica, foi proposta como molécula alvo em diferentes tipos de câncer.
Palavras-chave: Ceruloplasmina humana; Proteínas multifuncionais; Reagentes de fase aguda; Ferroxidase; Multicobre oxidase
La ceruloplasmina (CP) se describió hace casi 78 años; se encarga de transportar el 95% del cobre en el plasma sanguíneo y se aisló por primera vez en 1948 a partir de la fracción alfa-2-globulina sérica; se la reconoció como reactante de fase aguda y se le adjudicó su participación en la respuesta inflamatoria. Es una proteína que cataliza diferentes reacciones químicas. Se la ha identificado como una de las “proteínas azules” debido a que en su estructura molecular contiene seis iones cobre y presenta considerable homología de aminoácidos y similar cadena polipeptídica con la estructura denominada “cupredoxina”. En consecuencia, se la clasificó dentro de la familia de las multicobre oxidasas. Se ha visto que es primordial en el metabolismo del hierro, ya que puede actuar como oxidasa en la membrana del macrófago del sistema reticuloendotelial. El descubrimiento de su actividad peroxidasa le adjudicó la característica de multifuncionalidad que ha sido objeto de varios trabajos de revisión (1) (2) (3) .
En el presente artículo se abordarán algunos aspectos inherentes a su estructura molecular y a la relación de ésta como factor determinante de sus múltiples funciones. Además, se hará referencia a su participación en el metabolismo de iones y al concepto de multifuncionalidad. Luego se presentarán algunas consideraciones sobre la importancia de la determinación de CP en diferentes condiciones clínicas que podrían ser de gran utilidad en el seguimiento, pronóstico y evolución de individuos con o sin patología.
Las proteínas multifuncionales se definieron a finales del siglo XX. El término multifuncional se debe a Constance Jeffery, una bióloga estructural de la Universidad de Chicago y pionera en el campo de estudio de estas proteínas, quien lo ha difundido ampliamente y se refiere a la capacidad que presentan ciertas proteínas de realizar diferentes funciones bioquímicas o biofísicas según el medio biológico en el que se encuentren. Se han utilizado varios términos en la literatura para describir el fenómeno por el cual la misma proteína realiza dos o más funciones en el organismo (pleiotropía, proteínas multidominio, promiscuidad, proteínas multipropósito, etc.). Sin embargo, no han contribuido a la correcta definición de “proteína multifuncional” debido a que no existe consenso para su utilización porque el advenimiento de nuevos conocimientos la ha modificado sucesivamente (4) .
En la actualidad, el concepto de multifuncionalidad adquiere importancia a partir de los recientes hallazgos acerca del proteoma humano. Su identificación y conocimiento ha sido necesario para tratar de comprender a los sistemas biológicos involucrados en los procesos de salud y enfermedad. Se ha demostrado que contribuyen a su complejidad y robustez. También participan en la evolución de los organismos vivos porque pueden incrementar la capacidad funcional de un número limitado de genes o coordinar la comunicación entre vías metabólicas complejas de acuerdo con el tamaño del genoma. Por lo cual, la CP constituye un ejemplo de este tipo de proteínas ya que se sobrepuso al concepto paradigmático de que a partir de un gen se obtiene una única estructura molecular proteica con una función específica (5) .En la actualidad, el concepto de multifuncionalidad adquiere importancia a partir de los recientes hallazgos acerca del proteoma humano. Su identificación y conocimiento ha sido necesario para tratar de comprender a los sistemas biológicos involucrados en los procesos de salud y enfermedad. Se ha demostrado que contribuyen a su complejidad y robustez. También participan en la evolución de los organismos vivos porque pueden incrementar la capacidad funcional de un número limitado de genes o coordinar la comunicación entre vías metabólicas complejas de acuerdo con el tamaño del genoma. Por lo cual, la CP constituye un ejemplo de este tipo de proteínas ya que se sobrepuso al concepto paradigmático de que a partir de un gen se obtiene una única estructura molecular proteica con una función específica (5) .
La CP se encuentra formada por una cadena de 1046 restos de aminoácidos con un contenido de 7% a 8% de hidratos de carbono cuyo PM es de 132 kDa. Se halla codificada en 20 exones que abarcan 65 kb de ADN ubicados en el cromosoma 3q23-q24. El estudio de su estructura molecular por la técnica experimental de cristalografía por rayos X reveló que se encuentra formada por seis dominios constituidos por seis átomos de Cu, estrechamente unidos y dispuestos espacialmente de manera triangular. Se ha estudiado que tanto el Cu como el Fe son elementos que se hallan íntimamente relacionados a la molécula y a la función de la CP (6) .
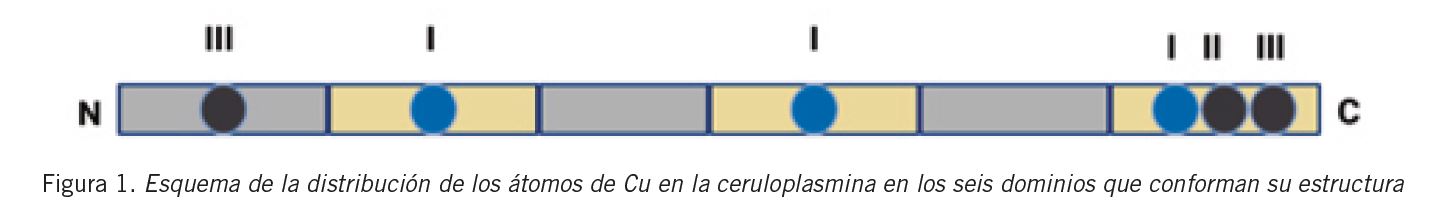
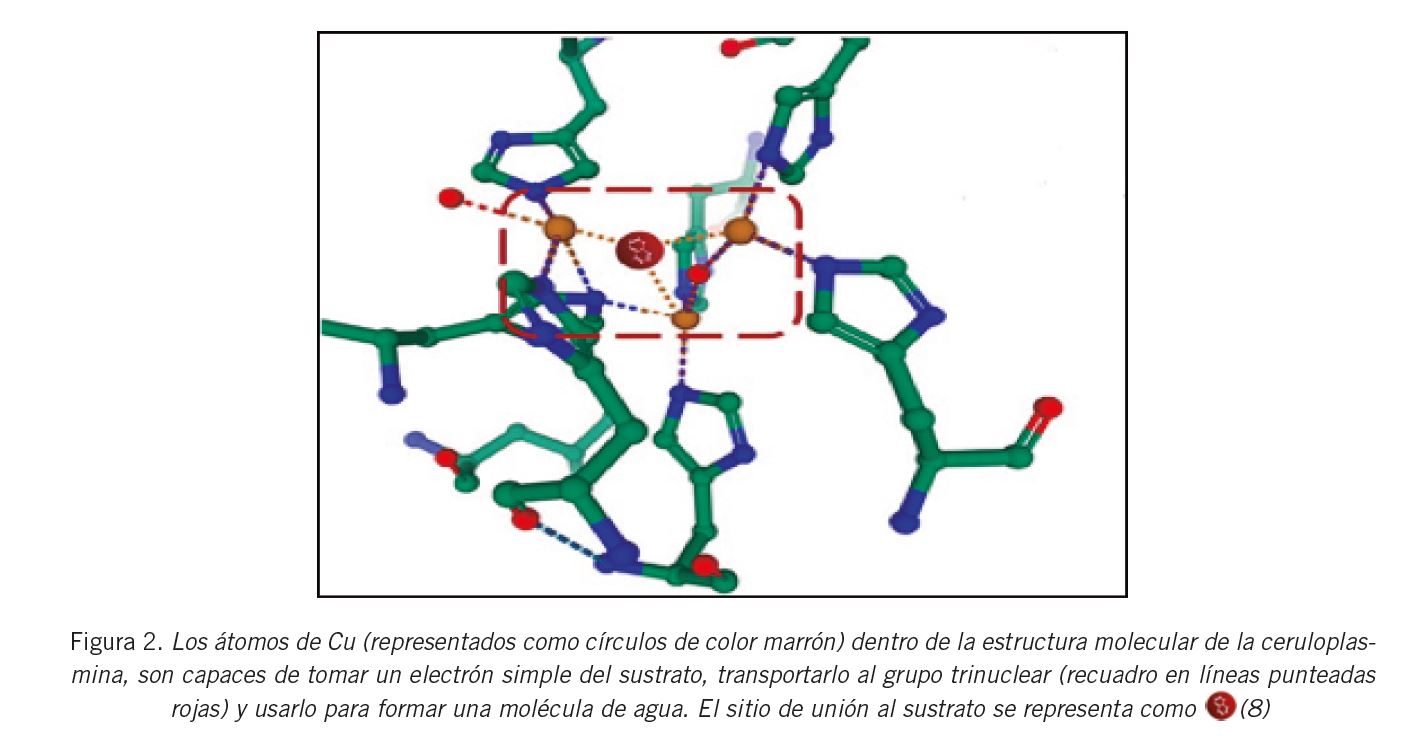
La conservación de la homología interna de la cadena polipeptídica constituye una de las características más relevantes de la estructura de la CP. Se encuentra formada por tres dominios homólogos a la cupredoxina, cada uno de ellos constituido por dos partes estructuralmente diferentes. Por lo tanto, la CP se halla integrada por seis dominios, los cuales se componen de tres tipos de iones Cu de acuerdo a sus características espectrofotométricas: el tipo I (T1Cu) con absorción a 610 nm (cobre azul); el tipo II Cu2+ (T2Cu) y el tipo III Cu2+ (T3Cu) (Fig. 1) . Este último, junto a los T2Cu no sólo participan en la actividad de la enzima, sino que también tienen la función de estabilizar a la estructura espacial mediante la formación de un cluster trinuclear (7) (Fig. 2) .
Los iones Cu se incorporan a la proteína durante el proceso de biosíntesis en el hígado (9) , que constituye la principal fuente de producción de CP. Además, se produce en otros órganos como los plexos coroideos del encéfalo, las glándulas mamarias, la placenta y el riñón (10) . Asimismo, los monocitos y los macrófagos la liberan a la circulación durante los procesos inflamatorios (11) .
En la circulación sanguínea, se ha demostrado que los iones Cu se pueden encontrar unidos fuerte o débilmente a la CP. Los primeros no son fácilmente liberados de su estructura y, para que esto suceda, existen dos posibilidades: en una se ha de producir la interacción de la CP con otra proteína y en la otra, con la superficie celular. Para los segundos tipos de unión, que son lábiles, se han identificado sitios de unión específicos, distintos de aquellos donde la unión es fuerte y pueden ser desplazados por otros iones como el Fe, el Co y el Ni (12) .
Se han descripto dos isoformas de CP producidas por splicing alternativo; una de ellas es secretada por el hígado y la otra es la que permanece unida a la membrana celular. En general, se ha visto que las especies extrahepáticas de la CP se encuentran unidas a la membrana celular a través del anclaje mediado por el grupo glicosilfosfatidilinositol (GPI), como se ha descripto en el cerebro de mamíferos (13) . Este tipo de unión también se expresa en el tejido hepático y en células inmunes, anclada a la membrana celular (14) .
Se ha demostrado que la CP humana es susceptible a la proteólisis por medio de enzimas proteolíticas como la trombina, in vitro durante la purificación o in vivo en los focos inflamatorios. En este sentido, se ha probado el rol protector de la CP en la artritis reumatoidea, ya que la trombina inhibe sus funciones anti-mieloperoxidasa y ferroxidasa, efecto que se revierte con la administración de hirudina, un potente inhibidor de la trombina (15) .
Con respecto a sus variantes genéticas, se ha investigado que pueden alterar tanto su expresión como sus funciones fisiológicas. Se ha visto que la CP se puede degradar rápidamente una vez liberada por el sistema reticuloendotelial, o bien puede permanecer dentro de éste con una estructura anormal impidiendo la incorporación de hierro a la misma y que se lleven a cabo sus funciones de oxidorreducción (16) .
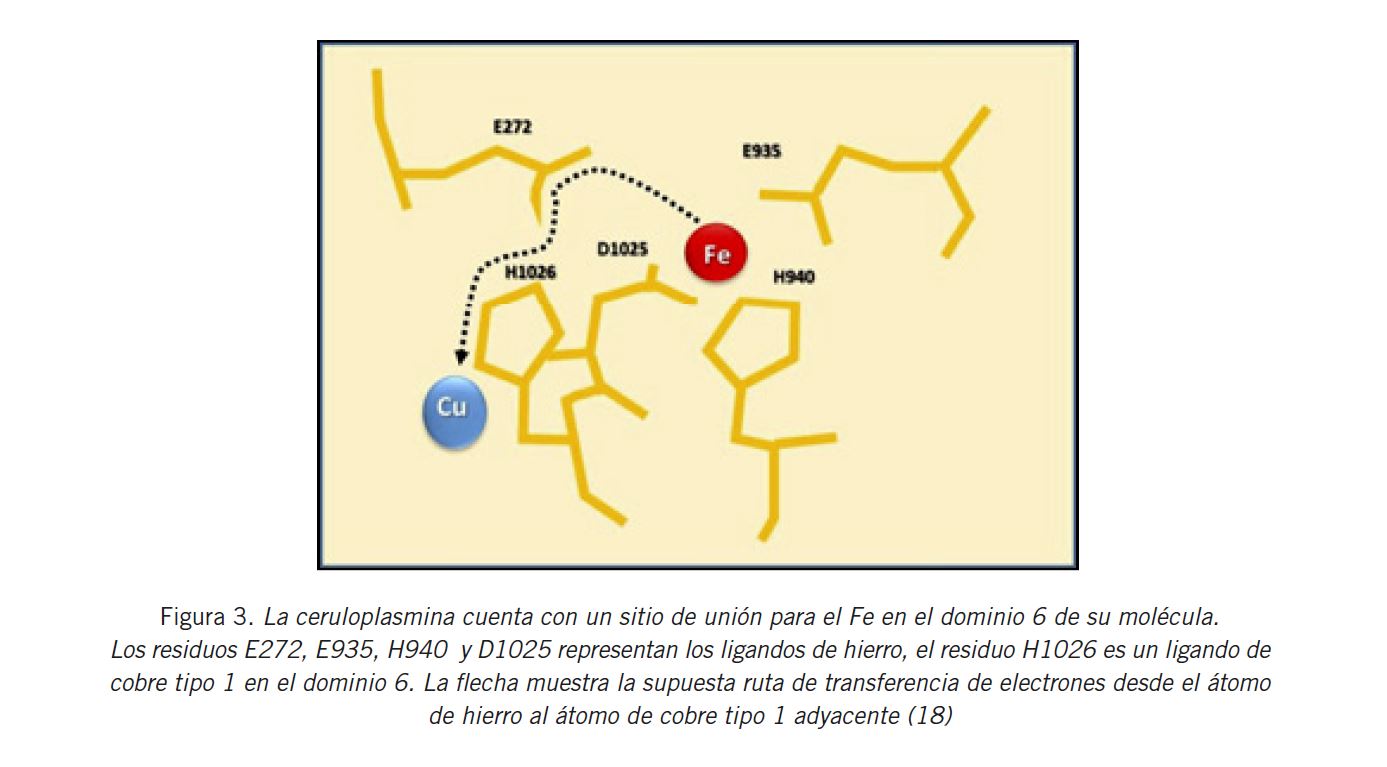
Por otro lado, se han descripto dos sitios de unión al Fe en los dominios 4 y 6 de la enzima (Fig. 3) . El acceso a estos sitios se encuentra permitido solo a sustratos relativamente pequeños, debido a la existencia de un impedimento estérico, en la parte superior de la molécula de CP. El Fe2+ y el Fe3+ parecerían unirse a los sitios llamados de retención, cercanos al lado exterior de la proteína. En el caso particular del Fe2+, esta situación podría explicarse porque se ha observado que el metal ocupa primero el sitio adicional o de oxidación de metales, que se ubica aproximadamente a 9,0–10,0 Å de distancia de los centros de Cu mononucleares respectivos y, por lo tanto, dentro del rango de transferencia de electrones. Allí es donde se produce la liberación de un electrón que es tomado por el Cu mononuclear más cercano. Posteriormente, sucede la translocación al sitio de retención con Glu 597 y Glu 935 en los dominios 4 y 6, respectivamente. Por consiguiente, se presume que cumple un papel clave en el proceso de translocación. En el sitio de retención, el Fe3+ se va a encontrar disponible para ser transportado, por ejemplo, por la transferrina (TF) sérica (17).
Las variantes alélicas de pérdida de función son responsables de la aceruloplasminemia, una condición hereditaria rara caracterizada por sobrecarga tisular de hierro, neurodegeneración y diabetes asociada, con hiperferritinemia y eritropoyesis deficiente en Fe (19). En individuos con hígado graso no alcohólico se observaron variantes genéticas que producen sobrecarga de hierro y metahemoglobinemia. En los portadores de estas variantes se observó fibrosis hepática más grave, lo que sugiere que la predisposición genética al depósito hepático de Fe puede traducirse en enfermedad hepática (14) .
Se observó que la formación de complejos da lugar a la modificación de los sitios de unión a los sustratos y puede alterar significativamente su función catalítica. Se ha visto que la CP se puede unir, interaccionar y formar complejos con metaloproteasas como la lactoferrina (LF) y la mieloperoxidasa (MPO). El modelo de complejo (CP: LF) se basó en el estudio por cristalografía por rayos X. En él se admitió que la estequiometría del complejo es 1:1, se demostró que ambos lóbulos de LF contactan con los dominios 1 y 6 de CP, se modifica el sitio de unión a las aminas biógenas y se impide su oxidación. Durante la fase aguda de la inflamación, los neutrófilos secretan LF como apoenzima (apo-LF) y se incrementa la síntesis de CP hepática, la cual se encarga de favorecer la incorporación de Fe a la LF y de generar su forma activa (holo-LF) (20) (21) (22) .
Se ha sugerido que la formación del complejo entre la CP y la MPO (2CP:1MPO) se encuentra relacionada con la interacción electrostática que se produce entre la naturaleza catiónica de la MPO y las cargas aniónicas de la CP. En consecuencia, la CP produce la inhibición de la actividad catalítica de la MPO y proporciona un escudo protector contra la generación excesiva de radicales libres, provenientes de la acción de peroxidación y clorinación de la MPO en estados inflamatorios. Por otro lado, la MPO protege las acciones antioxidantes de la CP al imposibilitar su clivaje entre los dominios 5 y 6 por la acción de las serino proteasas (tripsina, elastasa y plasmina). De esta manera, se preserva la integridad de la estructura molecular de la CP, requisito indispensable para la unión (2CP:1MPO). Se ha investigado la relación entre la actividad de la MPO en plasma, el nivel de CP y la actividad ferroxidasa I (FeOxI), junto con el estrés nitrosativo, los biomarcadores inflamatorios, neurohormonales y nutricionales. Se ha podido confirmar que en la insuficiencia cardíaca aumenta la actividad de la MPO y de la CP en plasma mientras que la de FeOxI disminuye, en comparación con individuos aparentemente sanos. Además, se ha demostrado una correlación positiva de la actividad de la MPO con la CP, mientras que no se ha encontrado entre la actividad de FeOxI y la MPO relacionada con la CP. El modelo del complejo ternario 2CP-2LF-MPO se ha descripto por dispersión de rayos X y se observó la ausencia de interacción entre la MPO y la LF. Se ha publicado que, junto con la MPO y la LF, algunas proteínas catiónicas de los leucocitos tales como la proteína catiónica eosinofílica, la catepsina G, la elastasa 3 neutrofílica y la azurocidina se pueden encontrar formando complejos con la CP (23) (24) (25) .
El conocimiento de las funciones fisiológicas de la CP se ha originado a partir de un gran número de investigaciones sobre la base del estudio de su estructura molecular, de su función ferroxidasa y de las propiedades físicas de los iones unidos a su molécula. Se ha visto que en respuesta a distintas condiciones celulares posee capacidad para ejercer diferentes funciones que dependen de la presencia de iones Cu en su molécula.
a. Actividad enzimática
Se han publicado numerosos trabajos que avalan su participación en diversas funciones fisiológicas, entre las cuales, la más importante es la de catalizar la reacción de oxidación del ión ferroso (26) (27) .
La nomenclatura de la CP se origina a partir del conocimiento de su actividad ferroxidasa. Los nombres por los que se la conoce son los siguientes:
La CP se puede unir a la molécula de oxígeno y generar agua como producto. Durante la catálisis, los electrones que provienen del sustrato son aceptados por el T1Cu y luego son entregados a los centros redox T2Cu / T3Cu del cluster trinuclear. Éstos se unen al oxígeno que se encuentra allí y se obtiene agua como producto de la reacción (28) . Se ha visto que en el curso del último paso de este proceso se pueden consumir y oxidar otros sustratos sin liberación de especies reactivas del oxígeno (ROS) (29) (Fig. 2) .
Al utilizar los iones Fe y Cu como sustratos puede catalizar la oxidación de Fe2+ a Fe3+ y la reducción de Cu2+ a Cu+, respectivamente. Además, actúa como aminooxidasa cuando oxida sustratos orgánicos como la fenilendiamina. La presencia de iones cloruro en concentración fisiológica estimula este tipo de reacción enzimática a pH ácido (30) . Se ha estudiado que puede oxidar a catecoles y sus análogos como la dopamina, epinefrina, norepinefrina, 5-hidroxitriptamina y el triptófano (31).
b. Actividad antioxidante
La acción antioxidante de la CP se ejerce a través de variados mecanismos. El más conocido se fundamenta en su actividad ferroxidasa. En general, se conoce que la oxidación de Fe2+ a Fe3+ se produce a través de la reacción de Fenton, y como productos de esta reacción se generan radicales hidroxilos y superóxidos, según se visualiza en la siguiente ecuación química:
En presencia de la CP, la oxidación del hierro se produce a través de su acción ferroxidasa, por lo que la reacción de Fenton no tiene lugar. De esta manera, se evita la formación de especies reactivas del oxígeno (ROS) y sus efectos deletéreos sobre las proteínas. Al comparar la capacidad scavenger que la CP y la superóxido dismutasa (SOD) ejercen sobre los radicales libres, se verificó que el efecto de la CP es más débil y relativamente constante (8). Su actividad antioxidante se demostró a través del estudio de individuos con aceruloplasminemia. En éstos se observó que la disminución de los niveles de CP y, en consecuencia, de su actividad ferroxidasa, se tradujo en una mayor disponibilidad de Fe2+ (32) .
Hacia fines del siglo XX se demostró que la CP inhibe la peroxidación lipídica, una reacción en cadena entre los ácidos grasos poliinsaturados y las ROS que genera peróxidos lipídicos y polímeros hidrocarbonados que son altamente tóxicos para la célula (33) (34) . También se ha demostrado que promueve la formación de S-nitroso glutatión y la expresión de óxido nítrico sintetasa (NOS) y, a través de este mecanismo, reduce el daño de los radicales hidroxilo en el tejido. Se ha visto que el óxido nítrico (NO) tiene un efecto citoprotector. La producción de RS-NO (nitrosotioles) en células HepG2 contribuyó a la citoprotección dependiente de NO e indujo la S-nitrosilación de varias moléculas que contienen tioles (SH) (35) . Recientemente se ha demostrado que existe asociación estadísticamente significativa entre los niveles elevados de lipoproteínas de baja densidad oxidada (LDLox) y su efecto prooxidante en individuos con diagnóstico de acromegalia activa (36) . Asimismo, se ha revelado que los niveles de CP se correlacionan negativamente con el valor de la escala unificada de valoración de la enfermedad de Parkinson (UPDRS) en la que se mide su severidad (37).
c. Metabolismo de iones
La CP participa en el metabolismo del Fe y en el transporte de iones Cu; ambos son elementos que se hallan involucrados en reacciones de oxidorreducción.
Se conoce que el Fe es el metal de transición más abundante en la naturaleza y se considera esencial en todas las formas de vida. En el ser humano se encarga del transporte y almacenamiento de oxígeno al formar parte del grupo prostético hemo, estructura fundamental de las proteínas hemoglobina, mioglobina, etc. Además, se encuentra involucrado en los fenómenos de oxidorreducción en los que participan citocromos, peroxidasas, catalasas, etc. (2).
Con respecto a su metabolismo, se sabe que el Fe no sólo forma parte constitutiva de la hemoglobina y de varias enzimas que son necesarias para el funcionamiento adecuado del organismo, sino también del citocromo P450 que participa de la detoxificación de drogas, de la síntesis de ADN, de la expresión génica y de la respuesta inmune. Se ha visto que los trastornos de su metabolismo pueden provocar obesidad e insulinorresistencia (38). El Fe en la dieta se encuentra en el máximo estado de oxidación (Fe3+), se reduce a Fe2+ por acción del citocromo B duodenal (DcytB) que se halla formando parte de la membrana superior de las células epiteliales intestinales. Luego, se ha de transportar a través de ellas por el transportador de metal divalente-1 (DMT1) y al unirse a la TF llega al hígado (39) (40). El transporte y la liberación de Fe a los distintos órganos se basa en la acción del sistema formado por la TF y el receptor de TF, principal transportador de Fe en humanos. El Fe se une a estas proteínas en su estado férrico (Fe3+).
En los últimos años se han identificado proteínas claves en el metabolismo del Fe, entre ellas la ferroportina (Fpt), una proteína que se encuentra ubicada en la membrana de los enterocitos y los macrófagos esplácnicos y hepáticos, entre otros tejidos. La CP contribuye a la estabilización de la Fpt en la membrana y se necesita que el Fe se encuentre en estado ferroso (Fe2+) para que se pueda unir a la Fpt (41). La oxidación del Fe en el hígado se encuentra mediada por la CP y en el enterocito por la hepaestina que es una proteína que tiene identidad con la CP (42).
Después de ingresar al hígado, el Fe se utiliza para sintetizar varias proteínas y el remanente se oxida (43). El transportador transmembrana requiere de la presencia en el hígado de Fpt, que es una proteína carrier específica. El sistema CP-Fpt es la principal vía de eflujo de Fe intracelular. Contrariamente a lo que sucede en el proceso de absorción de Fe en el que muchas proteínas se encuentran involucradas, el único sistema de excreción que se ha descubierto es el CP- Fpt (44).
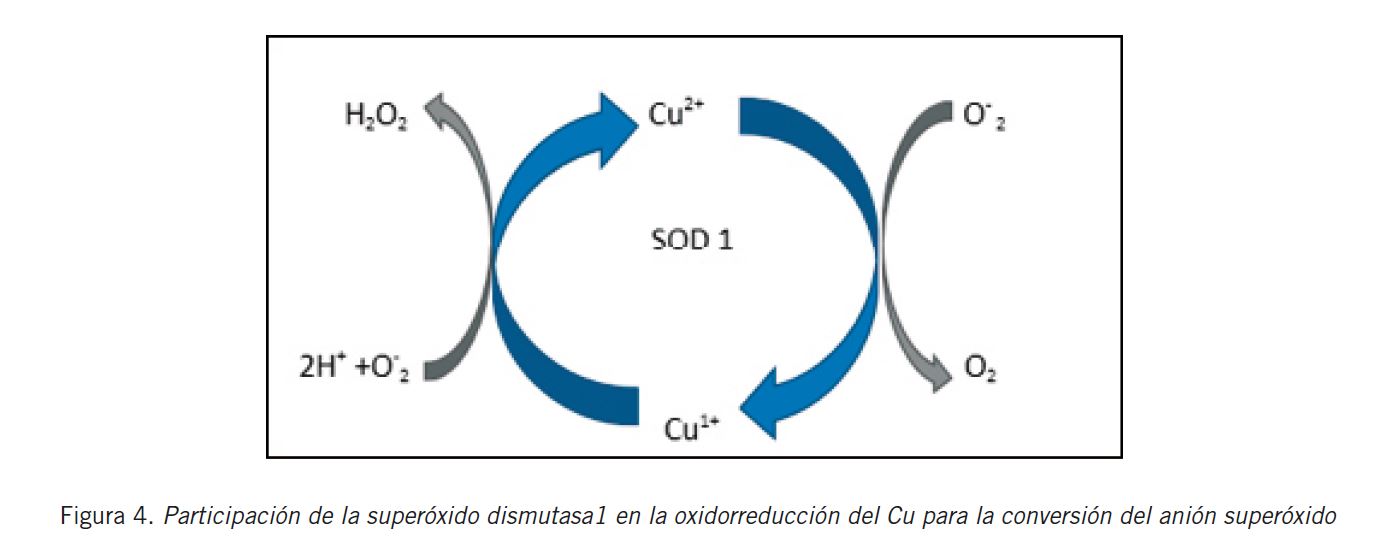
En referencia al Cu, se ha visto que se absorbe en el intestino delgado a través de la enzima ATP7A [proteína asociada al origen y desarrollo de la enfermedad de Menkes (45)], se une a la albúmina o a la α-2 macroglobulina para constituir el Cu no unido a la CP y se transporta hacia el hígado. Luego, se libera a los hepatocitos por medio de la proteína transportadora de Cu (CTR1) que se halla ubicada en la membrana celular. Este mecanismo de transporte no requiere de ATP. Dentro de la célula, cede el Cu a la enzima superóxido dismutasa 1 (SOD1) por acción de la chaperona de cobre (CCS). La localización de la SOD1 en el citoplasma, núcleo y espacio intermembrana de la mitocondria refleja la importancia de su actividad protectora en la célula. Esta actividad se realiza a través de la catálisis y dismutación del radical superóxido (O2 -) (Fig. 4).
A continuación, la chaperona de Cu de la citocromooxidasa (COX17) dona el Cu a las mitocondrias para sintetizar citocromooxidasa, la proteína 1 antioxidante (ATOX1) (34), que es la encargada de entregar el Cu a la ATP7B (proteína involucrada en la patogénesis de la enfermedad de Wilson) en la red trans-Golgi (Fig. 5).
Para finalizar, se incorpora el Cu dentro de la molécula de la CP y ésta se transforma en la principal transportadora de Cu. Cuando la CP llega a la superficie de la célula blanco, lo libera al interaccionar con receptores de superficie y se distribuye a los tejidos. La CP no unida al Cu se transforma en una proteína alostérica.
Independientemente de las funciones descriptas, el Cu es altamente tóxico, pues en su estado oxidado (Cu2+) participa en reacciones en las que se producen los radicales hidroxilo libres (OH-), el anión superóxido (O2 -) y el peróxido de hidrógeno (H2O2). Se ha observado que las ROS atacan a las biomembranas a través del proceso de peroxidación lipídica, desestabilizan su estructura y afectan sus funciones celulares (47).
d. Angiogénesis y cáncer
Se ha estudiado que la angiogénesis se produce en procesos fisiológicos y patológicos en el organismo vivo y que la formación de nuevos vasos sanguíneos requiere de la presencia de Cu y de elevadas concentraciones de CP en el sitio de crecimiento y de formación tumoral. Ésta, no solo aporta el Cu sino que también estimula el desarrollo de la neoangiogénesis, característica principal de las células malignas. Se ha observado la estrecha relación que existe entre la angiogénesis y la producción de ROS, probablemente debido a que las células cancerosas se encuentran en estrés oxidativo permanente ((48) (49).
Recientemente se determinaron los niveles de actividad de CP junto al factor de crecimiento endotelial vascular en una población de individuos con cáncer de ovario en diferentes estadios de la enfermedad y se concluyó que ambos presentaban valor pronóstico (50). Además, se observó que se encontraba asociada con el estadio T avanzado y la invasión perineural. Por eso, tiene la posibilidad de ser un marcador pronóstico candidato para el cáncer de las vías biliares (51). También se postuló a la CP en plasma como un biomarcador en el cáncer de células escamosas de la hipofaringe (52) (53). En contrapartida, la baja expresión de CP se correlaciona con un pronóstico favorable para el cáncer invasivo de mama (54). En terapias dirigidas que apuntan contra la CP se observó disminución de la angiogénesis y de la tumorogénesis en modelos de cáncer de colon (55)

La CP es una proteína multifuncional porque presenta la capacidad de ejercer diferentes y múltiples funciones tisulares en el organismo vivo. La conformación de su estructura molecular se modifica de acuerdo con las características de la célula y podría influir en la determinación de su función.
La CP posee la capacidad de unirse a proteínas y formar complejos binarios y ternarios. El complejo (CP:LF) podría considerarse un mecanismo de protección de la célula contra los productos de oxidación que se liberan por el estallido de la cadena respiratoria. Por otra parte, la importancia de la unión con la lactoferrina se debe a la regulación de la disponibilidad de Fe en las patologías hematológicas. En el caso del complejo 2CP:1MPO, se ha visto que puede ejercer su función antioxidante mediante la interacción proteína-proteína in vivo. Así, la interacción electrostática con la MPO impide la producción de radicales libres en exceso. Sin embargo, ésta se podría afectar frente a modificaciones postraduccionales de la estructura de la CP e implicaría el rol fundamental de la integridad de la proteína en la formación del complejo y su influencia en la función que desempeña. Se ha supuesto que la capacidad de la CP para formar complejos con las proteínas catiónicas leucocitarias podría ser parte de un mecanismo regulador en la patogénesis de la vasculitis sistémica (24). En el modelo del complejo ternario 2CP-2LF-MPO, la ausencia de interacción entre la MPO y la LF podría indicar que la CP actúa como mediador entre ambas moléculas proteicas (25).
En los procesos inflamatorios, las ROS podrían oxidar directamente a las proteínas, desnaturalizar el ADN y el ARN y provocar daños que contribuyan al desarrollo de diversas enfermedades como el cáncer, las enfermedades neurodegenerativas y al envejecimiento celular (47). La potencial actividad antioxidante de la CP protege a los tejidos de los efectos nocivos del Fe2+ al transformarlo en Fe3+ y al catalizar la producción de nitrosotioles (RS-NO). En el caso particular del GS-NO (nitrosoglutatión) actúa como neuroprotector (35). Particularmente, la disminución de los niveles de CP circulante se relaciona con el depósito de Fe en los tejidos. En el cerebro se estudió que promueve el desarrollo de enfermedades neurodegenerativas tales como el Alzheimer y otras, como la enfermedad de Parkinson o el autismo, debido al incremento de la peroxidación lipídica y a la generación de compuestos tóxicos.
Sobre la base de datos recientes se podría concluir que un nivel alto de actividad de CP podría ser útil para reconocer a las personas con mayor riesgo de enfermedad cardíaca crónica (ECC). La liberación sistémica de MPO que conduce a valores incrementados de su concentración sérica y la hiperceruloplasminemia podrían considerarse un rasgo característico de ECC asintomática e indicarían aterosclerosis en pacientes que eventualmente desarrollen ECC (23).
Asimismo, debido a su participación en los procesos de angiogénesis, hecho trascendental en el crecimiento, la proliferación y la metástasis de los tumores, se la podría considerar un blanco terapéutico en enfermedades oncológicas. La hipoxia que se genera en los procesos tumorales podría ser la causa de la estimulación de síntesis a través de la sobreexpresión del gen específico para CP en las células neoplásicas vía el factor 1 inducible por hipoxia (HIF-1), elemento indispensable para la adaptación a esta condición tisular. Además, en casos como el cáncer de mama se ha evidenciado su valor pronóstico al predecir la eficacia de la inmunoterapia. Su determinación podría ser de gran utilidad clínica para estratificar a estos pacientes y aplicarles el tratamiento farmacológico correspondiente.
El presente trabajo fue realizado sin haberse recibido una financiación específica.
Las autoras declaran no tener conflictos de intereses respecto del presente trabajo.
Bioq. VIVIANA MÓNICA YAPUR
Área Gastroenterología y Enzimología Clínica. Departamento de Bioquímica Clínica. Hospital de Clínicas “José de San Martín”. Facultad de Farmacia y Bioquímica. Universidad de Buenos Aires. (C1120AAR) Av. Córdoba 2351. Ciudad Autónoma de Buenos Aires, Argentina.
Correo electrónico: vmyapur@ffyb.uba.ar
Recibido: 24 de mayo de 2024
Aceptado: 26 de diciembre de 2024